
November 15, 2013 (Vol. 33, No. 20)
Live-Cell Microfluidic Platform Has Been Developed to Monitor the Entire Process
Autophagy leads to the lysosomal degradation of cytosolic components and organelles. The best understood role for autophagy is directing the removal of damaged or unwanted products. Autophagy can be induced by cancer therapies, when autophagy functions as a survival mechanism and potentially limits drug efficacy.
In established tumors, malignant progression and tumor maintenance have been linked to physiological adaptations resulting in upregulated or constitutively active autophagic pathways. Further, many stimuli can activate autophagy including nutrient starvation, reactive oxygen species, endoplasmic reticulum stress, and ammonia.
During autophagy, cytosolic proteins and aging organelles are sequestered by a double-membraned autophagosome. One of the hallmarks of autophagy is translocation of LC3 from the cytoplasm to the autophagosome. Autophagosomes then fuse with lysosomes, promoting breakdown of the vesicle and all contents, including LC3. This process can be visualized using a LC3-GFP fusion protein or an anti-LC3 antibody.
Many newer autophagy assays rely on the expression of stably transfected green fluorescence protein (GFP)-LC3 fusion proteins; autophagosome activity is identified by changes in GFP puncta.
Lysosomal inhibitors, such as chloroquine (CQ), have helped determine autophagic response to cellular stress. CQ blocks the last step of autophagy, lysosomal degradation; the resulting buildup of intermediates is a marker of autophagic activity. By combining the use of live-cell imaging with transduction of a GFP-tagged autophagosome marker (LC-3) in the presence of CQ, researchers can real-time monitor autophagosome formation on a fluorescent microscope. However, little is known about the latter stages of autophagy and lysosomal degradation.
Here, we used a microfluidic live-cell imaging platform (CellASIC™ ONIX Microfluidic Platform, EMD Millipore) to develop a dynamic cell-based assay for monitoring the entire autophagy process. This platform offers temperature and gas control, as well as media perfusion for precise environmental control. LC3-GFP CHO reporter cells were subjected to nutrient starvation or hypoxic stresses followed by reintroduction of normal growth conditions. With this approach, we have established a dynamic autophagy assay, which visualizes, under a fluorescent microscope, a real-time course of the autophagy mechanism.
The assay method described provides quantitative information on both autophagosome formation and lysosomal degradation machinery, providing a platform for the discovery of new targets and therapeutic compounds in cancer and other diseases.
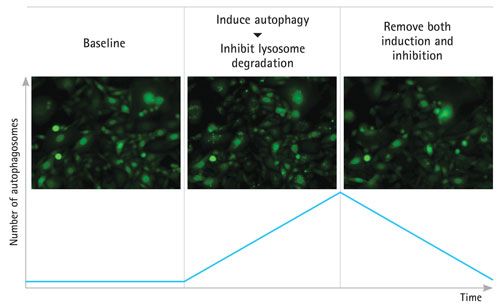
Figure 1. Schematic of live-cell imaging for autophagy of LC3-GFP–expressing CHO cells: Medium was perfused to establish fluorescent baseline. A stressor and the lysosome inhibitor CQ1 were introduced to trigger autophagosome accumulation. When cells were returned to standard growth medium, autophagosomes underwent lysosomal degradation.
Materials and Methods
Cell seeding in the CellASIC M04S Microfluidic Plate
A 1 x 106 cells/mL suspension was prepared, and the liquid in inlet wells 1, 6, 7, and 8 was aspirated. The inner ring in wells 6 and 7 was aspirated, and cell suspension was added into the inner ring of well 6. The plate was placed in a cell culture hood to allow cells to load into the microchambers by capillary action and incubated. Culture medium was added into well 1, and gravity-driven perfusion was used to deliver it to the microchamber.
Live-cell imaging for starvation-induced autophagy assay
Phosphate-buffered saline (PBS) was aspirated into wells 1, 2, 3, 6, 7, and 8, but the liquid in the inner holes was not disturbed. The desired treatments and culture medium were added into wells 2 and 3, respectively. The plate was sealed to the heater manifold and microincubator controller. The flow program was created using the CellASIC ONIX FG Software.
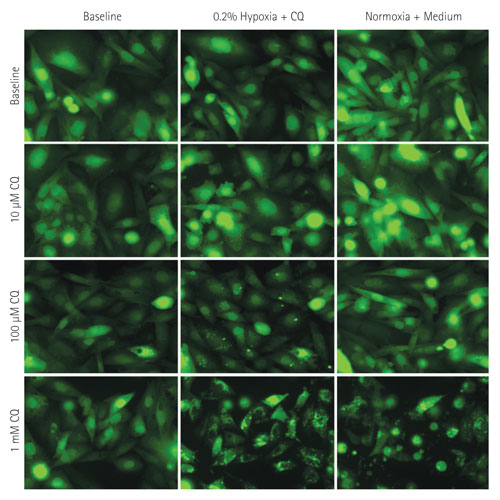
Figure 2. Images of the LC3-GFP CHO reporter cells taken during the hypoxia-induced autophagy assay: Cultures were perfused with standard growth medium under normoxic conditions, followed by continuous CQ perfusion in severe hypoxia treatment, followed by removal of the stressors and reestablishment of normoxia in standard growth medium. An Olympus® IX-71 inverted microscope was used; images were taken under the 40x objective.
Live-cell imaging for hypoxia-induced autophagy assay
The microincubator controller was connected to a premixed gas tank containing 95% air and 5% CO2 (normoxic gas line) and purged. The hypoxic gas line was also purged. PBS was aspirated from wells 1, 2, 3, 6, 7, and 8; liquid was left in the inner holes. The desired treatments and culture medium were added to wells 2 and 3, respectively. The flow program was created using the CellASIC ONIX FG Software.
Live-cell imaging and image analysis
An inverted microscope was used for live-cell imaging under the 40x objective. The number of autophagosomes was determined using an image processing sequence for object identification in CellProfiler Software (Broad Institute).
Results and Discussion
To validate the media exchange capability of the CellASIC ONIX platform and the ability to quantify autophagy through autophagosome counting, LC3-GFP CHO were perfused with EBSS + 50 µM CQ followed by regular culture medium. The dynamic changes of autophagy in both the stress and recovery phase could be quantified (Figure 1). Rate of autophagosome formation and degradation was successfully monitored.
Profiling of the CQ dose response in CHO cell lines was conducted. Once established in the microchamber, exposure conditions involved three phases: standard culture medium, continuous CQ perfusion (10 µM, 100 µM, or 1 mM), and culture medium. The rate of autophagosome formation was proportional to the CQ concentration. However, at 1 mM, cells ceased committing to the autophagy pathway; the number of autophagosomes stayed constant. We also observed more dead cells in this treated group, indicating either that maximal levels of autophagy had been achieved, or cells committed to apoptosis at the high CQ dose.
The CellASIC ONIX system was then used to introduce severely hypoxic conditions. Compared to the hypoxic response of cells cultured in traditional petri dishes, LC3-GFP CHO reporter cells in the microfluidic perfusion environment were far more sensitive to gas switching, demonstrating autophagosome formation within three hours of hypoxic treatment. Following a six-hour exposure, many cells underwent apoptosis.
Profiling of autophagosome formation in reporter cells in response to CQ under hypoxia conditions was performed, and the rate of autophagosome appearance accelerated with increasing CQ dose. As for the recovery phase, cells treated with 100 µM of CQ responded almost instantaneously, while those treated with the highest dose demonstrated a more protracted recovery profile (Figures 2 and 3).
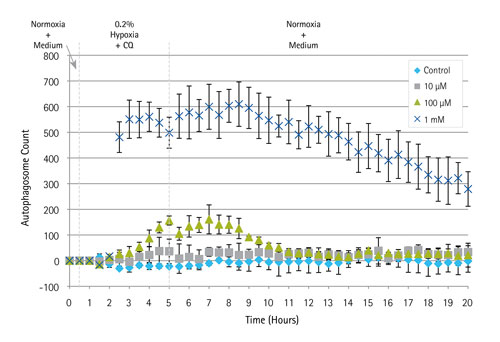
Figure 3. Quantitative live-cell imaging of autophagosome formation: Three levels of CQ inhibitor were perfused through independent culture units. Time-lapse imaging was performed on three different positions in each microchamber. The fluorescence intensity was counted and averaged per frame using CellProfiler software and normalized to the background to measure flux. Error bars represent S.D. of the puncta in around a total of 60 cells per time point.
Conclusion
The CellASIC ONIX platform was used to create an assay with the potential to simultaneously monitor multiple intracellular components throughout the entire autophagic process without disruption. It also allows precise manipulation of culture parameters, exposing cells to more physiologically relevant conditions.
The real-time analysis capabilities of this platform will provide new insights into how cancer cells may avoid therapeutic intervention and have large implications for the investigation of dynamic cellular processes.
Cindy Chen is R&D scientist, Terry Gaige is science and technology, engineer, Ivana Zubonja is R&D associate, and Paul Hung ([email protected]) is senior R&D manager at EMD Millipore.

