
June 1, 2015 (Vol. 35, No. 11)
Angela L. Huang, Ph.D.
Cellular Metabolism Live-Cell Assays for Research and Drug Discovery
Cellular energy production depends on mitochondrial integrity and function. Mitochondrial dysfunction has been shown to alter the cell cycle, metabolism, cell viability, gene regulation, and other key aspects of cellular growth and survival. In cancer studies, glycolysis persists to build ATP production for tumor growth while bypassing the need for healthy mitochondria (the Warburg Effect). Although the underlying genetic reasons for the causalities between aerobic glycolysis, tumor growth, and hypoxia remain unclear, there is evidence connecting cancer cells’ abilities to bypass normal cellular metabolic pathways to mitochondrial dysfunction.
Research from neurodegenerative disorders (e.g., Alzheimer’s, amyotrophic lateral sclerosis (ALS), Huntington’s, and Parkinson’s) has shown the importance of mitochondria in neuronal survival, cellular metabolism, and reactive oxygen species (ROS) production as well. Neurons depend on oxidative phosphorylation as a critical energy supplier and are very sensitive to ROS intracellularly. Thus, mitochondrial biogenesis and dysfunction have been linked to neurodegeneration and ageing.
Genetically encoded biosensors have proven to be powerful reporters of cellular activities and are highly amenable to customized designs to suit experimental needs. Their specifications can be fine-tuned to fluorescence range, kinetics, and cellular compartmental targeting, and can be incorporated into a variety of cell types. At Tempo Bioscience, we recently developed two biosensors, TempoATPTM and TempoO2TM, for cytotoxicity and cellular metabolism studies.
These biosensors can be incorporated into immortalized cancer cell lines, tumors, human inducible pluripotent stem cells (iPSCs), and iPSC-derived cell types such as neural stem cells, astrocytes, and other neuronal or glial cell types. Here, we report a new set of biosensor assays and show how they can be used for basic research, drug discovery, cytotoxicity studies, and chemical compound screening.
Genetically Encoded Intensiometric Biosensors
Intensiometric biosensors change fluorescence intensity when bound to an ion or molecule. Fluorescence intensity is also dependent on the biosensor concentration in each cell and on “thickness of the cell,” in addition to the ion concentration. Compared to traditional FRET-based biosensors that exhibit a shift in the absorption or emission spectra, intensiometric biosensors offer the following advantages: 1) higher sensitivity (larger dynamic range), 2) smaller spectral bandwidth (avoiding high background noise levels), and 3) a requirement for only one set of fluorescence filters. Traditional FRET reporters are limited in number of donor/acceptor pairings by fluorophore requirements. Most fluorophores have broad excitation-to-emission spectrums, making energy transfer prone to signal-to-noise concerns. In comparison, intensiometric imaging is quantitative and thus less prone to artifacts.
Genetically encoded biosensors, in contrast to chemically synthesized molecules such as Fura2/3/4, are made by the cell and become functional without further intervention by the scientist, simplifying assay procedures. In addition, chemical dyes/sensors demonstrate difficulties targeting to specific cellular organelles and tend to become compartmentalized at random, before getting extruded from the cell during lengthy recording experiments.
Genetically based biosensors can be more precisely and elegantly targeted, using rational genomics and structural designs. Targeting to subcellular compartments, such as the mitochondrial targeting of TempoMitoTM, which measures calcium fluctuations in the mitochondria, is a significant advantage of genetically encoded biosensors.
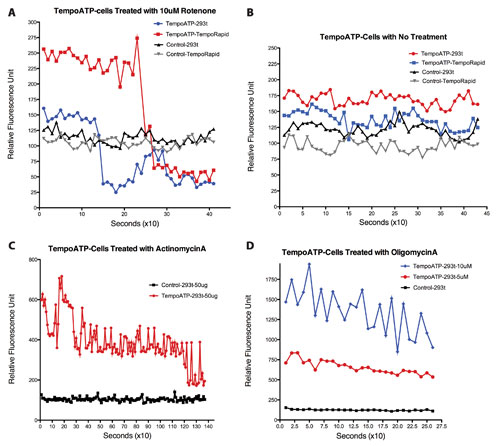
Figure 1. (A) Response to 10 µM rotenone in TempoATP-incorporated HEK293t and TempoRapid™ cells monitored over time (seconds). Representative traces shown (n = 7). (B) Baseline responses from HEK293t and TempoRapid cells without treatment over time (seconds). Representative traces shown (n = 5). (C) Responses from TempoATP-incorporated HEK293t cells to actinomycin A treatment (50 µg). Representative traces shown (n = 12). (D) Oligomycin A treatment in TempoATP-incorporated HEK293t cells (seconds; 5 µM vs. 10 µM). Representative traces shown (n = 12).
Experimental Need
Cellular metabolism can be measured by a variety of compounds, molecules, and ions as byproducts of a complex set of pathways that are required for cells’ survival and growth. Examples are: lipid metabolism, proteasome degradation, and carbohydrate metabolism. ATP and oxygen are critical indications of aerobic cellular respiration and oxidative phosphorylation, and the aerobic pathway depends on mitochondria, a critical organelle in the cell. Thus, mitochondrial dysfunction is a vital indication of cytotoxicity.
Not surprisingly, mitochondrial retention of ATP molecules only constitutes ~25% of overall cellular respiration and leaked oxygen due to ROS-induced stress accumulates in the cytosol. Thus, cytosolic measurements of ATP and O2 can determine the effects of internal or external factors on cytotoxicity in most cases, although targeting measurements to the mitochondrial inner membrane can be useful under special circumstances.
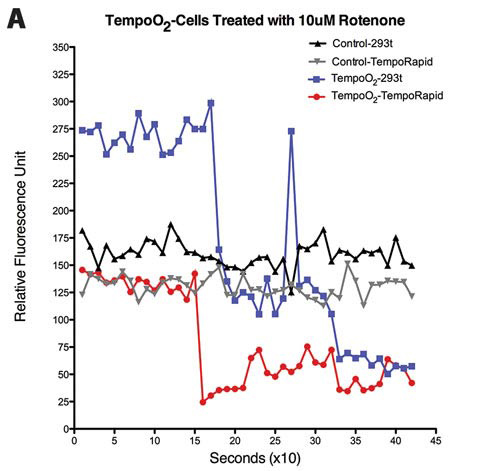
Figure 2. (A) Response to 10 µM rotenone in TempoO2-incorporated HEK293t and Tempo-RapidTM cells monitored over time (seconds). Representative traces shown (n = 7).
Cytosolic ATP and O2 Biosensors
Incorporated into cells, TempoATP reports cellular fluctuations in ATP. Figures 1A–1D show cellular responses to various mitochondrial inhibitors, rotenone, actinomycin A, and oligomycin A at 617 nm (excitation)/635 nm (emission).
Similarly, TempoO2 reports cellular fluctuations in O2, which partly depends on the mitochondria also. Figures 2A–2C show cellular responses to various mitochondrial inhibitors, rotenone, actinomycin A, and oligomycin A at 617 ± 2 nm (excitation)/635 nm (emission).
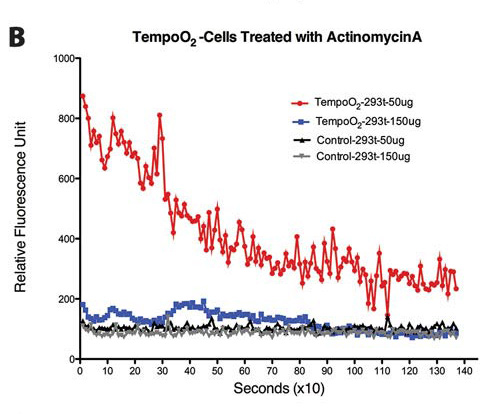
Figure 2. (B) Responses from TempoO2-incorporated HEK293t and TempoRapid cells to actinomycin A treatment (50 µg vs. 150 µg). Representative traces shown (n = 12).
Conclusion
TempoATP and TempoO2 are biosensors incorporated into specific cell types of interest for basic research, preclinical drug discovery, and cellular toxicity studies. They are functional reporters for the activation or inhibition of cellular respiration and mitochondrial function. They can be incorporated into target-specific and phenotypic assays.
At Tempo Bioscience, we believe that genetically encoded intensiometric biosensor technologies provide a wide range of assay opportunities for scientists performing research and functional studies related to preclinical, cytotoxicity, and drug development studies in the global biopharmaceutical industry.
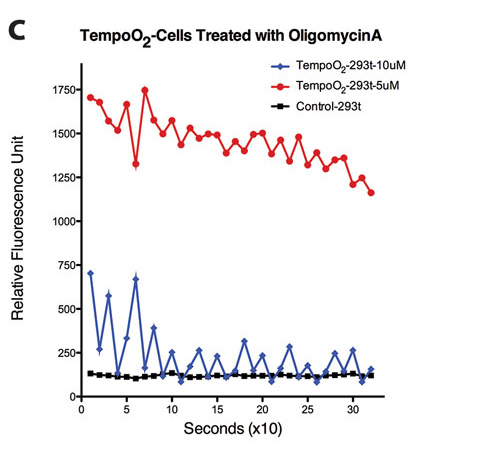
Figure 2. (C) Oligomycin A treatment in TempoO2-incorporated HEK293t cells (seconds; 5 µM vs. 10 µM). Representative traces shown (n = 12).
Angela L. Huang, Ph.D. (angela@tempo bioscience.com), is founder of Tempo Bioscience.





